
成長相転換(花成)の制御〜その3〜
1.5.2 FT遺伝子 - 花成の促進遺伝子 -
機能喪失変異体の表現型、発現制御の様態から、FT遺伝子は、長日条件下でCO遺伝子依存的にその転写が活性化され、花成を促進する遺伝子であると考えることができる。このことから、FT遺伝子を構成的に発現させた場合には、光周期・CO遺伝子機能の有無とは無関係に、早熟花成が引き起こされることが期待される。実際に、FT遺伝子を構成的に高発現させた形質転換植物を作出してみると、期待された表現型が得られた(図7)(Kobayashi et al. 1999)。
一方、CO遺伝子の構成的な高発現によっても光周期に依存しない早熟花成が引き起こされることが報告されていたが、FT遺伝子の機能欠損変異がこの早熟花成を強く抑圧することが最近になって報告されている。
FT遺伝子が "光周期依存促進経路" の制御下にあることは、これらの結果から明らかであるが、一方で、短日条件下、あるいはCO遺伝子の機能欠損下においてもFT遺伝子の発現は消失しない(図6C)。このことは、FT遺伝子の発現が "光周期に依存しない経路" によっても制御されていることを示している。そのような経路として、FCA遺伝子が関与する "自律促進経路" が考えられる(図3)。実際、最近になって、生育条件によっては、FT遺伝子の発現がFCA遺伝子の機能欠損変異において顕著に減少することを確認している(基礎生物学研究所 中村研三教授・大藤雅章博士との共同研究。Ohto et al. 2001)。また、FT遺伝子を構成的に発現させた形質転換植物の早熟花成表現型がFCA遺伝子の機能欠損変異の影響をほとんど受けないことも確認している(小林恭士・荒木 崇, 未発表)。
そうした知見に加えて、"春化依存促進経路"のFRIGIDA (FRI)遺伝子、"自律促進経路", "春化依存促進経路" の共通の制御ターゲットである花成抑制遺伝子FLCとの遺伝学的関係(小林恭士・荒木 崇, 未発表)から、FT遺伝子はこれら2つの経路の下流にも位置づけられることが明らかになってきた。
われわれのグループを含む複数のグループの研究から、FT遺伝子, 1.4 節で言及したLFY遺伝子, そして、SUPPRESSOR OF OVEREXPRESSION OF CO 1(SOC1. AGAMOUS-LIKE 20(AGL20)ともいう)遺伝子の少なくとも3つの遺伝子が花成を制御する4つの経路を統合するという図式が浮かび上がりつつある(図8)(Araki 2001)。
これら3つの遺伝子の活性を合わせたものが、花成のいわば"スイッチ"にあたることは、これら3つのうちの2つ(FT遺伝子とLFY遺伝子)を構成的に高発現させた形質転換植物が発芽後、栄養成長を経ることなく直ちに花成することによって端的に示されている(図9)。

図7 FT遺伝子を構成的に高発現させた形質転換シロイヌナズナ
(A) 非形質転換植物(WT Columbia, 左)とFT遺伝子を構成的に高発現させた形質転換植物(p35S::FT, 右。
(B) CiFT遺伝子を構成的に高発現させた形質転換植物(p35S::CiFT)
(C) TSF遺伝子を構成的に高発現させた形質転換植物(p35S::TSF)

図7 FT遺伝子を構成的に高発現させた形質転換シロイヌナズナ(つづき)
(D) 形質転換植物個体の全体像。
2枚のロゼット葉(矢尻)と2枚の茎葉(矢印)を形成し、花成した個体。茎頂分裂組織は頂花(TF)に転換している。スケールは 5mm。
(E) 茎頂部分の拡大像。
3個の花が融合した頂花に転換している。スケールは 2.5mm。
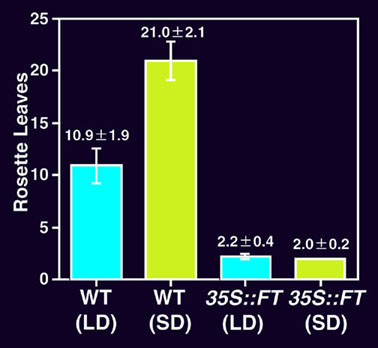
図7 FT遺伝子を構成的に高発現させた形質転換シロイヌナズナ(つづき)
(F) 長日(LD, 16時間明・8時間暗)と短日(SD, 8時間明・16時間暗)条件における表現型の比較。
非形質転換植物(WT)とFT遺伝子を構成的に高発現させた形質転換植物(35S::FT)の表現型を、形成されたロゼット葉の数で比較した。
非形質転換植物では、2つの条件で花成までの時間には顕著な差が見られる。これに対し、形質転換植物では、2つの条件下で有意差はなく、花成の光周期反応性が失われている
。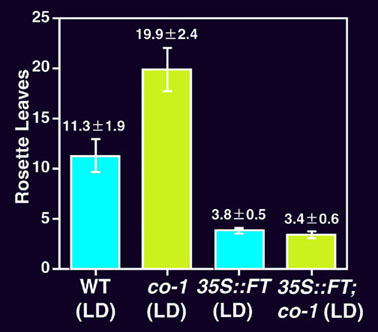
図7 FT遺伝子を構成的に高発現させた形質転換シロイヌナズナ(つづき)
(F) co-1変異の有無による表現型の比較。
非形質転換植物(WT)とFT遺伝子を構成的に高発現させた形質転換植物(35S::FT)のそれぞれで、co-1変異の有無による表現型の差を、形成されたロゼット葉の数で比較した。長日(LD, 16時間明・8時間暗)条件における表現型。
非形質転換植物では、co-1により、花成は顕著に遅れる。これに対し、形質転換植物では、co-1変異の有無による有意差はなく、co-1変異の表現型は完全に抑圧されている。
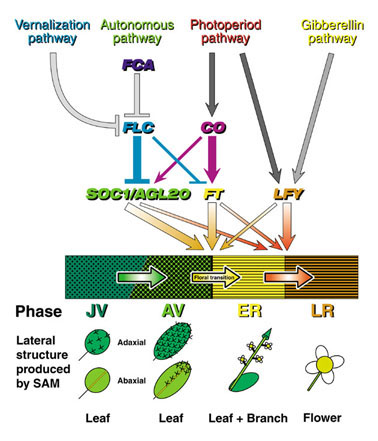
図8 FT遺伝子と花成を制御する経路の統合
シロイヌナズナの後胚発生過程(中央の横長の帯で示す)は、幼若期栄養成長相(JV)、成熟期栄養成長相(AV)、初期生殖成長相(ER)、後期生殖成長相(LR)の4相に分けることができる。AV から ER への移行(中央の黄色の矢印)が花成である。各相において茎頂分裂組織が形成する側生構造をそれぞれの下に模式的に示す。
JV と AV では葉が形成されるが、形状、毛(表皮細胞が変形したもの)の分布がこの二つの相では異なる。ER では茎葉伴う側枝(花序)が、LR では花が、それぞれ形成される。
花成の制御に関わる4つの経路は、SOC1/AGL20, FT, LFY の3つの遺伝子の発現制御を介して統合される。それぞれの経路の相対的な寄与は3つの遺伝子で異なる(矢印とT字バーの太さで表現)。また、花成とER から LR への移行に対する寄与も3つの遺伝子で異なる(矢印の太さで表現)。FT遺伝子の発現制御に最も大きく寄与するのは、CO遺伝子を介した光周期依存経路であり、FT遺伝子は花成を主に制御し、ER から LR への移行には小さな寄与しか持たない。
See Curr. Opin. Plant Biol. 4(1), 63-68 (2001).

図9 FT, LFY遺伝子を同時に構成的に高発現させた形質転換植物
非形質転換植物(WT Columbia, 左)とFT, LFY遺伝子を同時に構成的に高発現させた形質転換植物(p35S::FT; p35S::LFY, 右。別個体植物の走査型電子顕微鏡像(下図)へ)。
それぞれの右側には植物体の模式図を示す。非形質転換植物では、2枚の子葉(薄黄色の楕円で示す)の間に数枚の本葉(緑色の楕円で示す)が形成され、栄養成長をおこなっているが、形質転換植物では、2枚の子葉の間に1つの花が形成され、全成長を終える。
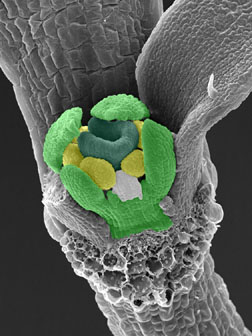
FT, LFY遺伝子を同時に構成的に高発現させた形質転換植物(p35S::FT; p35S::LFY)。
子葉展開直後の植物の茎頂分裂組織を示す。萼片(薄緑色)、花弁(白色)、雄蘂(黄色)、心皮(緑色)の4種類の花葉器官原基がみとめられる。奥に見えるのは子葉。手前の子葉は切除してある(切断面が三日月状に見えている)。この個体では2枚の抱葉が形成されている(左側の1枚を切除している)。